Жизненный цикл ВИЧ связан только с определенными клетками человека, в окружающей среде без человека вирус быстро погибает.
Общая схема поведения вируса в клетках изображена на рис. 8 . В целом эта схема сходна у всех ретровирусов, отличия только в деталях, но именно эти детали и делают ВИЧ смертельно опасным.
Все вирусы для того, чтобы поразить организм, прикрепляются к клеткам хозяина, связываясь со специфическими белками — рецепторами. Рецепторы для разных типов вирусов различны ( табл. 2 ). Этим предопределяется, какие клетки могут быть заражены (инфицированы) данным вирусом, а какие нет. Так, рецептор для вируса полиомиелита имеется только на нейронах; они инфицируются в первую очередь при попадании вируса в организм. Риновирус соединяется с белком ICAM-1 , который присутствует на мембранах многих типов клеток, в результате все они могут могут быть заражены этим вирусом. На сегодняшний день не для всех вирусов обнаружены рецепторы.
ВИЧ, попав на слизистую оболочку или в кровяное русло человека ищет то место, куда мог бы проникнуть и где мог бы существовать, чтобы в дальнейшем размножиться. ВИЧ способен проникать не во все виды клеток крови, а только в те, которые несут на своей поверхности специальный белок-рецептор — CD4 , к которому вирус легко присоединяется ( табл. 2 ). Белок вируса gp120 ( рис. 6 ), расположенный на его поверхности, находит белок-рецептор CD4 на поверхности клетки и плотно связывается с ним по принципу «ключ-замок». Этому взаимодействию способствуют дополнительные белки — корецепторы — CCR5 и CXCR4 . В клетках они служат рецепторами для специфических клеточных белков-регуляторов — хемокинов. А для взаимодействия клеток с ВИЧ они играют роль помощников для основного рецептора CD4. Без этих белков-корецепторов, также как без CD4- рецептора, вирус проникнуть в клетку не может.
Рецепторы и корецепторы для ВИЧ имеются на поверхности нескольких типов клеток иммунной системы. Наличие CD4-рецептора позволяет называть все эти клетки CD4-лимфоцитами. В частности, на мембране Т-лимфоцитов- T-хелперов имеется CD4-рецептор и CXCR4-корецептор. CD4-рецептор содержится также на поверхности макрофагов и дендритных клеток , которые одновременно несут и корецептор CCR5 ( рис. 9 ). На ранней стадии ВИЧ- инфекции вирусы обычно имеют большее сродство к макрофагам, поэтому их называют M-тропными . Белок оболочки этих вирусов gp120 способен связываться одновременно с CD4-рецептором и CCR5-корецептором. На более поздних стадиях ВИЧ приобретает сродство к T-лимфоцитам , поскольку белок gp120 видоизменяется и становится способным связываться с клетками, содержащими как CD4-рецептор, так и CXCR4-корецептор. По этой причине такие вирусы называют T-тропными .
CD4-белок-рецептор — важный клеточный белок, который обычно участвует на самых первых этапах сложного процесса передачи сигналов при активации Т-клеток. ВИЧ сумел подобрать «ключ» именно к этому «замку». В результате в организме человека атака вируса идет главным образом именно на CD4-содержащие клетки. Основным способом попадания ВИЧ внутрь таких клеток человека является его физическое связывание как с белком- рецептором, так и белком-корецептором, расположенными на клеточной оболочке ( рис. 9 ).
Происходящее за этим слияние внешней оболочки вируса с мембраной клетки-мишени обеспечивает легкое проникновение (перетекание) вируса внутрь клетки. При этом ВИЧ полностью освобождается от своей оболочки.
Медицинский эксперт статьи

Возбудитель ВИЧ-инфекции
Описано два серотипа — ВИЧ-1 и ВИЧ-2, различающиеся по структурным и антигенным характеристикам. На территории Украины эпидемиологическое значение имеет ВИЧ-1 (основной возбудитель заболевания).
Систематика ВИЧ
Вирус иммунодефицита человека относят к царству Viridae, семейству Retroviridae, подсемейству Lentiviridae.
Основные свойства вируса иммунодефицита человека
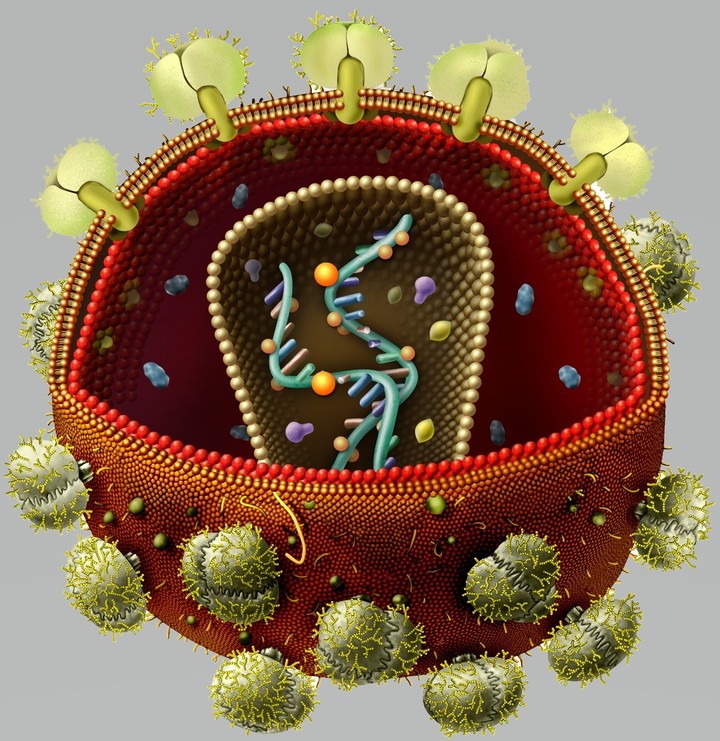
По данным электронной микроскопии, вирус имеет округлую форму и сложную структуру. Диаметр вириона составляет 100-120 нм.
Группы белков ВИЧ-1 и ВИЧ-2
Белки оболочки вируса (env)
gp160, gp120, gp41
gр140, gр105, gр36
Белки сердцевины (gag)
Ферменты вируса (pol)
Молекулярный вес белков измеряют в килодальтонах (кДа): gp — гликопротеины; р — протеины.
В центре вириона располагается геном вируса, представленный двумя нитями РНК, внутренними протеинами р7 и р9, а также ферментами — обратной транскриптазой (ревертазой), протеазой, РНКазой и интегразой (эндонуклеазой). Геном окружает внутренняя белковая оболочка. В состав внутренней оболочки ВИЧ-1 входят протеины р17, р24 и р55. Протеины р16, р25 и р56 образуют внутреннюю оболочку ВИЧ-2. Наружная липидная оболочка ВИЧ-1 пронизана гликопротеином gpl60. состоящим из трансмембранного (gp41) и высокоиммуногенного (gpl20) фрагмента. Оболочечные белки gp41 и gpl20 соединены нековалентной связью и формируют на поверхности вириона отростки, обеспечивающие присоединение ВИЧ к рецепторам клеток-мишеней человека.
Геном вируса содержит девять генов — три структурных и шесть регуляторных. Геном является изменчивой структурой за счёт антигенного дрейфа. Существует ряд серологических вариантов вируса (например, А, В, С, D, E, F, G, Н).

[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13], [14], [15]
Устойчивость вируса в окружающей среде
В естественных условиях ВИЧ (в высушенном состоянии) сохраняет активность в течение нескольких часов; в жидкостях, содержащих большое количество вирусных частиц, таких, как кровь и эякулят, — в течение нескольких дней.
В замороженной сыворотке крови активность вируса определяют на протяжении нескольких лет.
Нагревание до 56 °С в течение 30 мин приводит к снижению инфекционного титра вируса в 100 раз. При более высокой температуре (70-80 °С) вирус погибает через 10 мин. При обработке вирионов 70% раствором этилового спирта в течение минуты происходит их инактивация. При воздействии 0.5% раствора гипохлорита натрия, 1% раствора глутаральдегида, 6% раствора пероксида водорода, 5% раствора лизола, эфира или ацетона также отмечают гибель вирусных частиц.
ВИЧ относительно мало чувствителен к ультрафиолетовому облучению и ионизирующей радиации.
Патогенез ВИЧ-инфекции
Дифференцировочный антиген CD4+ (CD — аббревиатура от Cell Differention antigen) и неспецифические (независящие от наличия CD4+) компоненты — рецепторы для ВИЧ. CD4+ — гликопротеид с молекулярной массой 55 000 кД. по своему строению схожий с определёнными участками иммуноглобулинов. Аналогичное строение имеет и белок вируса gpl20 — это и определяет способность проникновения вирионов в клетку.
Рецептор CD4+, расположенный на мембране иммунокомпетентных клеток, выполняет функцию распознавания антигенов (вместе с белками HLA — главного комплекса гистосовместимости II класса).
Типы клеток, поражаемых вирусом иммунодефицита человека
Фолликулярные дендритные клетки
Толстый кишечник, почки
Клетки шейки матки
Оболочка ВИЧ содержит белки человеческого комплекса гистосовместимости I и II классов, поэтому проникновение вируса в организм не вызывает реакции отторжения. Фиксация вирионов на поверхности клетки-мишени происходит при участии гликопротеина gpl20. Гликопротеин gp41 обеспечивает слияние вирусной оболочки с мембраной клетки-мишени. Двуцепочечная РНК вируса проникает в клетку, где с помощью обратной транскриптазы происходит синтез одноцепочечной провирусной ДНК. Затем происходит формирование двуцепочечной ДНК, встраивающейся в ДНК клетки с помощью интегразы. Вирусная ДНК выполняет функцию матрицы для синтеза РНК, собирающей новую вирусную частицу.
Нередко при репликации ВИЧ возникают генетические ошибки. Таким образом происходит образование разных подтипов вируса.
После проникновения ВИЧ в СD4+-клетки начинается его репликация: чем активнее СD4+-клетки, тем интенсивнее репродукция вируса. Поэтому регуляторы, активирующие CD4+-клетки обеспечивают увеличение репликации вируса. К подобным регуляторам относят ФНО: фактор, стимулирующий колонии гранулоцитов и макрофагов (колониестимулирующий фактор), и ИЛ-6.
Интерферон и трансформирующий фактор роста — регуляторы, тормозящие репликацию вируса. Как показали исследования. ФНО-а активирует транскрипцию провирусной ДНК ВИЧ-1 в хронически инфицированных Т-клетках и макрофагах. Моноциты, синтезирующие ФНО-а. не только индуцируют экспрессию ВИЧ-инфицированными клетками, но и стимулируют активацию латентного провируса. Регистрируют одновременную активность ФНО-а. ИЛ-6 и фактора, стимулирующего колонии гранулоцитов и макрофагов.
Иммунопатогенетические признаки ВИЧ-инфекции — дефицит Т-звена и В-звена иммунной системы: недостаток компонентов комплемента и фагоцитов; снижение функций неспецифических факторов защиты. Поликлональная активация В-лимфоцитов приводит, с одной стороны, к гипергаммаглобулинемии, а с другой — к ослаблению способности клеток продуцировать вируснейтрализующие антитела. Происходит увеличение количества ЦИК и образование антител к лимфоцитам; это вызывает ещё большее уменьшение числа СD4+-Т-лимфоцитов. Отмечают развитие аутоиммунных процессов. Поражение иммунной системы при ВИЧ-инфекции носит системный характер.
Наряду с дефицитом СD4+-лимфоцитов в течение болезни нарастает функциональная недостаточность СD8+-лимфоцитов, NK-клеток (естественных киллеров) и нейтрофилов. При ухудшении иммунного статуса развиваются различные инфекционные, аллергические, аутоиммунные и лимфопролиферативные заболевания, а также синдром, свойственный болезни иммунных комплексов (эти факторы определяют клиническую картину ВИЧ-инфекции).
На начальных стадиях заболевания в организме вырабатываются вируснейтрализующие антитела, подавляющие активность свободно циркулирующих вирусов. Однако такие антитела не действуют на вирусы, находящиеся в клетках (провирусы). С течением времени (обычно после 5-6 лет) происходит истощение защитных возможностей иммунной системы и, следовательно, накопление вируса в крови.
После заражения ВИЧ проникает в лимфоидные клетки Лангерганса, которые локализуются в слизистой и в коже, затем инфицируются макрофаги и CD4+-лимфоциты, с которыми вирус распространяется по всему организму. Клетками-мишенями ВИЧ являются Т-лимфоциты, дендритные лимфоциты и клетки Лангерганса (их незрелые предшественники), моноциты/макрофаги, эозинофилы, мегакариоциты, тимоциты, некоторые клоны В-лимфоцитов, клетки нервной системы — нейроны, микроглиальные клетки/макрофаги. Все эти клетки объединяет наличие на их мембране рецептора СD4* (* от англ. Cell Differentiation Antigen 4 — дифференцировочный клеточный антиген № 4. Номер соответствует порядковому номеру очередного препарата (ОКТ-4) серии моноклональны.х антител, полученных фирмой «ORTHO» (США) для исследования рецепторов лимфоцитов), к которому имеет большое сродство вирусный эпимембранный гликопротеин gp120 (ВИЧ-1) и gp105 (ВИЧ-2) [25].
Вместе с тем следует заметить, что существует ряд клеток, которые, не имея рецептора СD4, селективно сорбируют, транспортируют на мембране или проводят через себя ВИЧ. К такого типа клеткам относятся М-клетки слизистой прямой кишки, граничащие с лимфоидной тканью стенки кишки, и сперматозоиды.
Из перечисленных клеток-мишеней, несущих на мембране CD4, важнейшими в патогенетическом плане являются Т-лимфоциты, выполняющие функцию хелперов/индукторов и эффекторов, их количество является наибольшим среди клеток, несущих рецептор CD4, в системе крови человека (табл. 9.1.)
| Таблица 9.1. Прямые клетки-мишени для ВИЧ в системе крови человека, имеющие на мембране рецептор CD4 | |
| Тип дифференцированных клеток | Содержание в крови. %* (норма) |
Т-лимфоциты:
|
|
| Дендритные лимфоциты | 1 |
| Моноциты/макрофаги | 3-1 |
| Эозинофилы | 0,5-5 |
| В-лимфоциты (некоторые клоны) | 1,8-8,5 |
| * от числа ядросодержащнх клеток крози. ** в основном несут на мембране рецептор СD8. |
|
По тропизму изоляты ВИЧ делят на моноцитотропные и лимфоцитотропные. Первые преобладают на начальных стадиях болезни, вторые — в период разгара.
Процесс взаимодействия ВИЧ с клеткой-мишенью включает ряд последовательных стадий. Проникновению вируса внутрь клетки предшествует взаимодействие оболочечных белков вируса с молекулами, экспрессированными на мембране клетки. Необходимым условием фиксации ВИЧ на клеточной мембране и последующего проникновения его в клетку является не только наличие рецептора CD4, с которым соединяется белок gp120 (ВИЧ-1) или gp105 (ВИЧ-2), но и корецептора. Для макрофаготропной разновидности ВИЧ таковым является хемокиновый рецептор ССR5, для лимфоцитотроп-ой — СХСR4.
Хемокиновый рецептор обеспечивает конформационное изменение трансмембранпого белка gр41 (ВИЧ-1) и gр36 (ВИЧ-2), необходимое для слияния вируса с клеточной мембраной. Некоторые варианты ВИЧ могут использовать оба корецептора.
У 2% населения на макрофагах отсутствует хемокиновый рецептор ССR5 вследствие генетических мутаций, что обусловливает невосприимчивость таких людей к ВИЧ-инфекции.
На следующей стадии оболочка зафиксированного на клеточной мембране вириона сливается с клеточной мембраной, сердцевина освобождается от оболочки (стадия «раздевания») и проникает в клетку. При этом вполне возможно, что вирусная оболочка становится частью наружной мембраны клетки-хозяина. Проникшая в цитоплазму свободная сердцевина вируса раскрывается. Далее с помощью обратной транскриптазы по матрице вирусной геномной РНК осуществляется синтез ДНК, для чего используются азотистые основания из цитоплазмы. Молекулы вирусной ДНК приобретают кольцевидную форму (циклизация) и проникают из цитоплазмы в ядро, где встраиваются (интеграция) в состав гепома клетки-хозяина [16].
Таким образом, процесс инфицирования вирусом иммунодефицита человека клетки-мишени можно разделить на следующие стадии:
- Связывание вириона с поверхностью клетки. Рецепция вируса.
- Слияние мембран вируса и клетки. Проникновение вируса внутрь клетки.
- Высвобождение нуклеоида и геномной РНК вируса.
- Синтез провирусной ДНК по матрице геномной РНК вируса.
- Интеграция генома провируса в геном клетки.
- Активация процесса транскрипции с ДНК провируса, трансляция белков вируса.
- Активная репликация вируса, то есть продукция всех компонентов вируса и формирование из них зрелых дочерних вирионов.
- Высвобождение вирионов и отдельных белков ВИЧ из клетки-хозяина во внешнюю среду и беспрепятственное заражение других клеток. Цитопатогенные эффекты ВИЧ.
Ведущим звеном в патогенезе ВИЧ-инфекции является поражение Т-хелперов [19], которое обусловлено:
- преждевременным старением и гибелью инфицированных клеток
- уничтожением зараженных клеток лимфоцитами-эффекторами антителозависимой клеточной цитотоксичности;
- блокадой рецепторов CD4 вирусным гликопротеином gр120;
- аутоиммунными процессами.
На Т-хелперы ВИЧ оказывает прямой цитопатогенный эффект. Истощение пула Т-хелперов приводит к тому, что они не могут полноценно обеспечивать функцию и взаимодействие других иммунокомпетентных клеток. Однако и на ранних этапах течения ВИЧ-инфекции, когда еще нет выраженного снижения содержания CD4+-клеток, а доля инфицированных CD4+-лимфоцитов не превышает 0,01% их числа [21], основное значение в развитии дисбаланса иммунного ответа, формировании иммунодефицита принадлежит нарушениям регуллториых функций Т-хелперов/индукторов |2б|. Причиной указанных нарушений является блокада рецептора СD4.
Распознавание Т-хелперами антигенов на поверхности антигенпрезептирующих клеток (например, макрофагов) происходит с обязательным участием двух структур — рецептора CD4, который взаимодействует с главным комплексом гистосовместпмостп II класса (ГКГС II) макрофага, и рецептора для антигена. Связывание только молекулы CD4 каким-либо лигандом (например, анти-CD4-антителом или вирусным белком gp120) в отсутствие последующего задействования рецептора для антигена является для клетки «отрицательным сигналом». При этом происходит быстрая и существенная элиминация молекул CD4 с поверхности клетки. В результате такой лимфоцит не может нормально взаимодействовать с белками главного комплекса гистосовместимости II класса (ГКГС II), находящегося на антигенпрезентирующих клетках, что является необходимым условием для формирования нормального иммунного ответа.
Взаимодействие поверхностного гликопротеина gр120 с мембраной CD4+-клеток может не только вызывать негативные сигналы, но и приводит к программированной клеточной гибели — апоптозу зрелых CD4+-лимфоцитов или гематопоэтнческих СD34+-клеток-предшественников даже при отсутствии инфицирования клеток ВИЧ.
G. Furlini с соавт. [18] установили, что спустя три часа после воздействия ВИЧ-1 (или очищенного рекомбинаитпого белка gp120) на CD4+-клетки in vitro наблюдается пик увеличения синтеза и ядерной транслокации белков теплового шока семейства БТШ-70. Эти данные свидетельствуют о способности gp120 запускать каскад процессов, используя сигнальную активность мембран. Одним из таких внутриклеточных процессов является активация системы белков теплового шока, что в свою очередь указывает на нахождение клетки в неблагоприятных условиях и формирование клеточной стресс-реакции.
Таким образом, не только полноценные вирусы, прямо инфицирующие Т-лимфоциты-хелпeры, но и отдельный растворимый вирусный белок gp120 вызывают иммуносупрессию путем связывания молекул CD4. Особенно значительный иммуносупрессорный эффект оказывает агрегированный (например, специфическими антителами) белок gp120 [26].
Гомология аминокислотных последовательностей молекул вирусного белка gp120 и рецепторпых структур клеток (HLА класса II и CD4) обусловливает появление перекрестно реагирующих антител (аутоантител), которые нарушают кооперативные взаимодействия иммунокомпетентных клеток. Аутоиммунная реакция формируется и в отношении иммуноглобулинов, под структуру которых мимикрирует gp120.
Нарушение функции Т-хелперов влечет за собой снижение цитотоксической активности Т-супрессоров, на которые ВНЧ также оказывает цитопатогенный эффект, что, в свою очередь, обусловливает появление оппортунистических инфекции и СПИД-индикаторных опухолей.
С развитием выраженной виремии количество инфицированных клеток в крови и интенсивность их гибели возрастает. От момента инфицирования до терминальной стадии СПИД содержание CD4+-клеток уменьшается более чем в 20 раз.
В моноцитах/макрофагах, в отличие от Т-лимфоцитов, ВИЧ реплицируется с умеренной интенсивностью, вирионы оформляются в округлые частицы еще в цитоплазме клетки и, выходя из нее, не оказывают цитонекротического действия. Однако моноциты/макрофаги, инфицированные ВИЧ, хотя и не подвергаются быстро наступающему цитолизу при массовом высвобождении дочерних вирионов, как Т-лимфоциты, но претерпевают значительные ультраструктурные преобразования. Указанные изменения, видимо, являются причиной пониженной бактерицидной (фунгицидной) активности и способности к хемотаксису моноцитов и макрофагов, а также ослабления взаимодействия их рецепторов с Fс-фрагментами иммуноглобулинов.
В-лимфоциты, учитывая отсутствие или невысокую плотность CD4-реиепторов на их цитоплазматнческой мембране, должны повреждаться в меньшей степени. Косвенным подтверждением этого, казалось бы, служит тот факт, что суммарная концентрация иммуноглобулинов IgG и IgА сыворотки в условиях ВИЧ-инфекции оказывается повышенной. Однако у больных отмечается характерная диспропорция уровней подклассов иммуноглобулинов IgG. Так, показано, что содержание IgG1 и IgG3 у таких пациентов увеличено, тогда как концентрация IgG2 и IgG4 существенно уменьшена. Прогрессирующее снижение уровня IgG2 может объяснить возрастающую восприимчивость больных ВИЧ-инфекцией к патогенному действию таких микроорганизмов, как Haemophilus influenzae, Streptococcus pneumoniae и Staphylococcus aureus.
В-лимфоциты на фоне активной секреции антител характеризуются слабой реакцией на митогeны и на неоантигены. Таким образом, несмотря на гипергаммаглобулинемию, функциональное состояние В-системы иммунитета у больных ВИЧ-инфекцией сходно с состоянием, развивающимся на фоне выраженной гипогаммаглобулинемни [4].
Функционирование В-лимфоцитов контролируется Т-лимфоиитами. Поэтому, вероятно, дисфункции В-системы в целом могут быть вторичными по отношению к дисфункции Т-хелперов и Т-супрессоров. Кроме того, ВИЧ способен прямо инфицировать В-лимфоциты и вызывать их разрушение. Опыт показывает, что содержание В-лимфоцитов в периферической крови больных в стадии СПИД может быть в три с лишним раза ниже нормы.
Вторым объектом воздействия ВИЧ после иммунной системы является ЦНС. ВИЧ инфицирует нейроны, астроциты, микроглиальные клетки (макрофаги), эндотелий кровеносных сосудов, фибробластоподобные клетки мозга. Однако прямого цитопатогенного эффекта вируса на указанные клетки не отмечено. Поражение ЦНС, проявляющееся деменцией и другими неврологическими нарушениями при ВИЧ-инфекции, скорее связано с нейротоксическим действием gp120 и появлением аутоантптел против антигенов мозговой ткани [15].
Таким образом, прогрессированию течения ВИЧ-инфекции предшествует активация репликативного процесса вируса иммунодефицита человека. Клинические проявления заболевания обусловлены непосредственным патогенным эффектом вируса и его белков на клетки-мишени, истощением пула CD4+-клеток крови, а также нарушением кооперативных связей и функций иммунокомпетентных клеток, что приводит к формированию иммунодефицита.
| Страница 1 2 3 4 5 6 7 8 | всего страниц: 8 |
- Федеральный закон РФ «О предупреждении распространения в Российской Федерации заболевания, вызываемого вирусом иммунодефицита человека» от 30 марта 1995 года.
- Змушко Е. И., Белозеров Е. С. ВИЧ-инфекция / Руководство для врачей. — СПб: Питер, 2000. — 320 с.
- Исаков В. А., Аспель Ю. В., Богоявленский Г. В. и др. Опыт применения циклоферона в терапии ВИЧ-инфекцни и СПИДа / Руководство для врачей.- СПб, 1997.- 60 с.
- Кожемякин Л. А., Бондаренко И. Г., Тяптин А. А. Синдром приобретенного иммунодефицита / Пособие для врачей.- Л.: Знание, 1990.- 112 с.
- Лобзин Ю. В., Казанцев А. П. Руководство по инфекционным болезням. — СПб., 1996. — 712 с.
- Лысенко А. Я., Турьянов М. X., Лaвдовская М. В., Подольский В.М. ВИЧ-инфекция и СПИД-ассоциированные заболевания / Монография.- М.: ТОО «Рарогъ», 1996,- 624 с.
- Новохатский Л. С., Хлябич Г. Н. Теория и практика лабораторной диагностики синдрома приобретенного иммунодефицита (СПИД). — М.: ВИНИТИ, 1992,- 221 с.
- Покровский В. И., Покровский В. В. СПИД: синдром приобретенного иммунодефицита.- М.: Медицина, 1988.- 43 с.
- Покровский В. И. ВИЧ-инфекция или СПИД // Терапевт, арх. — 1989. — Т. 61, № 11. — С. 3-6.
- Покровский В. В. ВИЧ-инфекция: клиника, диагностика / Под общ. ред. В. В. Покровского.- М.: ГЭОТАР МЕДИЦИНА, 2000.- 496 с.
- Рахманова А. Г. ВИЧ-инфекция (клиника и лечение).- СПб: «ССЗ», 2000.- 367 с.
- Рекомендации по применению антиретровирусных препаратов у взрослых и подростков, инфицированных вирусом иммунодефицита человека // Consilium Medicum приложение. Январь 2000,- 22 с.
- Смольская Т. Т., Ленинская П. П., Шилова Э.А. Серологическая диагностика ВИЧ-инфекции / Методическое пособие для врачей.- СПб, 1992.- 80 с.
- Смольскал Т. Т. Второе десятилетие жизни в условиях СППДа: уроки и проблемы / Актовая речь.- СПб., 1997.- 56 с.
- Хаитов Р.М., Игнатьева Г. А. СПИД.- М., 1992.- 352 с.
- Connor S. Research shows how HIV exhausts the body // Brit. Mod. J.- 1995.- Vol.310.- P. 6973-7145.
- Burcham J., Marmor M., Dubin N. et al. CD4 is the best predictor of development of AIDS in a cohort of HIV-infecteci homosexual men // J. AIDS.- 1991.- jN»9. — P.365.
- Furlini G., Vignoli M., Re M. C., Gibellini D., Ramazzotti E., Zauli G.. La Placa M. Human immunodeficiency virus type I interaction with the membrane of CD4+ cells induces the synthesis and nuclear translocation of 70K heat shock protein // J.Gen. Virol.- 1994.- Vol.75, pt 1.- P. 193-199.
- Gallo R. C. Mechanism of disease induction by HIV // J.AIDS.- 1990.- N3.- P. 380-389.
- Gottlieb M. S., Schroff R., Schanker H. et al. Pneumocystis carinii pneumonia and mucosal candidiasis in previously homosexual mon // Now England J. Med. — 1981. — Vol. 305. — P. 1425-1430.
- Harper M. E., Marselle L. M., Gallo R.C., Wong-Staal F. Detection of lymfocytes expressing human T-lymphotropic virus type III in limph nodes and peripheral blood from infected individuals by in situ hybridization // Proc. Natl. Acad. Sci. U. S. A. — 1986. — Vol. 83. — N 2. — P. 772-776.
- Hess G. Clinical and diagnostic aspects of HIV-infection.- Mannheim: Boehringer Mannheim GmbH, 1992.- 37 p.
- Hu D.J., Dondero T.J., Ryefild M. A. et al. The emerging genetic diversity of HIV // JAMA.- 1996. — N 1.- P. 210-216.
- Lambin P., Desjobert H., Debbia M. et al. Serum neopterin and beta-2-microglobulin in anti-HIV positive blood donors // Lancet.- 1986.- Vol.8517. — P. 1216.
- Maldonado I. A., Retru A. Diagnosis of pediatric HIV disease // The AIDS knowledge base, Fd. Cohen P.T.; Sande M. A. Voiberding. 1994.- P. 8.2.1-8.2.10.
- Mc Dougal J.S., Kennedy M.S., Sligh J.M. et al. Binding of the HTLV-III/LAV to T4+ T cells by a complex of the 110K molecule and the T4 molecule // Science.- 1985.- Vol.23.- P. 382-385.
- Montagnier L., Gougeon M. L., Olivier R. et al. Factors and mechanisms of AIDS pathogenesis // Science challenging AIDS. Basel: Karger, 1992.- P. 51-70.
- Paterlini P., Lallemant-Le C., Lallemant M. et al. Polimerase chain reaction for studies of mother to child transmission of HIV-I in Africa // J.Med. Virol. — 1990.- Vol.30, N 10.- P. 53-57.
- Polis M. A., Masur H. Predicting the progression to AIDS // Amor. J. Med. — 1990.- Vol.89, N 6.- P. 701-705.
- Roddy M.M., Grieco M. H. Elevated soluble IL-2 receptor levels in serum of HIV infected populations // AIDS Res. Hum. Retrovir. — 1988.- Vol.4, N 2. — P. 115-120.
- Van dor Groen. G., Van Kerckhoven I. et al. Упрощенный и менее дорогой, в сравнении с традиционным, способ подтверждения инфекции ВИЧ // Бюлл. ВОЗ.- 1991.- Т. 69, №6.- С. 81-86.
Источник: Медицинская лабораторная диагностика, программы и алгоритмы. Под ред. проф. Карпищенко А.И., СПб, Интермедика, 2001
Вирусы могут размножаться только внутри клетки-хозяина, перестраивая ее работу так, что клетка начинает производить новые вирусные частицы. Это серьезно нарушает работу самой клетки и в большинстве случаев ведет к ее гибели. Процесс проникновения и размножения вируса в ней сложный и содержит много стадий, а значит, специальные лекарства могут тормозить его на различных этапах.
В случае с ВИЧ это выглядит так.
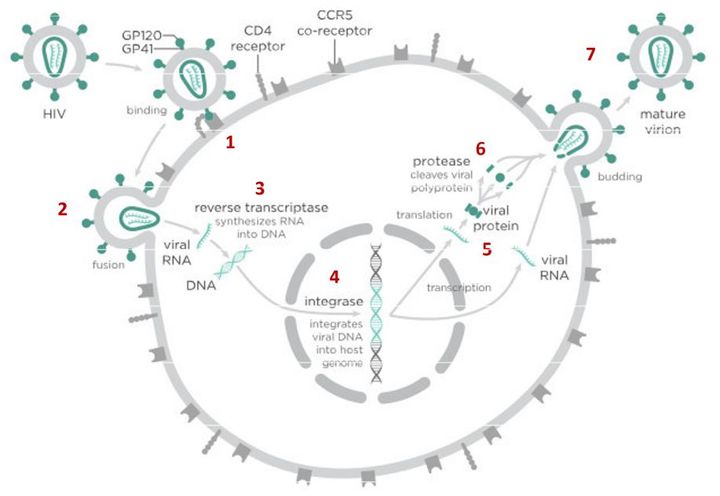

После того как вирусная частица приклеилась к рецепторам CD4 и CCR5/CXCR4 и закрепилась на поверхности клетки, другой специальный белок вируса gp41 соединяет оболочку вируса с мембраной клетки-мишени и помогает вирусу проникнуть внутрь клетки (цифра 2 на схеме). На этом этапе остановить вирус может другая группа препаратов — ингибиторы слияния, например, энфувиртид. Препарат связывается с вирусным белком gp41 и нарушает его работу.
На этом этапе размножение вируса блокирует очень большое количество препаратов из двух классов — нуклеозидные ингибиторы обратной транскриптазы (НИОТ — ламивудин, абакавир, тенофовир) и ненуклеозидные ингибиторы обратной транскриптазы (ННИОТ — эфавиренз, рилпивирин). Принцип действия у веществ из обеих групп похожий: молекула лекарства связывается с обратной транскриптазой вируса, нарушает ее работу и так блокирует перенос генов вируса из РНК в ДНК. Это не позволяет генам вируса встроиться в ДНК клетки-хозяина и начать производство новых вирусных частиц.
На следующем этапе инфицирования (цифра 4 на схеме) готовая вирусная ДНК проникает в ядро и встраивается в ДНК клетки-хозяина с помощью специального вирусного фермента — интегразы. Работу этого фермента блокируют ингибиторы интегразы, например, долутегравир, не позволяя ДНК вируса встроиться в наши гены.
После того как вирусная ДНК встраивается в ДНК клетки-хозяина, клетка получает неправильную программу и начинает производить вирусные белки и РНК, необходимые для сборки новых частиц вируса (цифра 5 на схеме), используя для этого структуры, которые в норме производят собственные белки клетки. Такая перенастройка производства белка сильно нарушает работу клетки и через некоторое время вызывает ее гибель. Лекарств, которые бы влияли на этот этап размножения вируса, нет, так как здесь вирус использует собственные ферментные системы клетки-хозяина и блокирование их работы какими-то веществами навредит здоровым клеткам.

После синтеза вирусных белков клеткой-хозяином в работу вступает еще один фермент вируса — протеаза (цифра 6 на схеме). Дело в том, что белки ВИЧ синтезируются в виде длинных цепочек белков-предшественников, но они неактивные. Протеаза разрезает эти цепочки на более короткие фрагменты, которые становятся активными белками вируса: из одних собирается вирусная оболочка, другие выполняют функции ферментов (обратная транскриптаза, интеграза, протеаза, о которых мы говорили выше). Класс лекарств, нарушающих работу вирусной протеазы, включает большое количество препаратов от достаточно старого лопинавира до современного дарунавира. Основной недостаток препаратов этого класса — высокая степень перекрестной устойчивости: если вирус приобретает устойчивость к одному препарату, скорее всего, на него не подействует ни один другой препарат из этого класса.
На последнем этапе (цифра 7 на схеме) вирусные частицы собираются из готовых компонентов (синтезированных клеткой вирусных белков и РНК), как конструктор, и покидают клетку-хозяина. Лекарств, которые бы тормозили эти процессы, пока не существует.
ВИЧ имеет высокую склонность к мутациям, что позволяет ему быстро приобретать устойчивость к лекарствам. Мутации изменяют форму молекул ферментов вируса. Если при этом форма изменится существенно, то фермент не сможет нормально работать и такой вирус размножаться перестанет — это вредная для вируса мутация. Однако в некоторых случаях — при полезных мутациях — форма молекулы фермента меняется не сильно, а так, что сам фермент продолжает нормально работать, но молекула лекарства уже не может с ним связаться.
Поэтому для эффективного подавления размножения ВИЧ обычно назначают три разных препарата из разных групп, влияющих на разные ферменты вируса, блокируя его воспроизводство на разных этапах. Исключением является группа ингибиторов обратной транскриптазы (НИОТ и ННИОТ) — эти лекарства связываются с разными участками молекулы вирусного фермента, поэтому могут назначаться вместе, несмотря на то, что относятся к одной группе. В условиях такой терапии вирусу очень трудно выработать устойчивость, потому что одновременно должно произойти несколько полезных мутаций в разных генах вируса, а это бывает крайне редко.
Есть несколько перспективных технологий, например, CRISPR/Cas9, которые могут позволить полностью удалить вирусную ДНК из организма человека, но пока ни одна из них не прошла все необходимые исследования для подтверждения эффективности и безопасности. Исследования продолжаются.